I know, I know. When you hear me say that biological sex isn’t real, you hear that I’m saying that the sexed attributes of a body aren’t real, that having sex isn’t real, and that reproduction isn’t real. But here’s the problem: you’ve conflated a map for the terrain.
To be clear: bodies and reproduction are material realities; your thoughts about that material reality are just that: thoughts.
The pop media term “biological sex” can mean only two things: the study of living organisms or the life processes of living organisms. To say “biological sex” is to reference the process category of an organism, which is to say, it’s ontology. Ontology is a branch of science that concerns itself with the nature of being. It strives to establish a set of concepts and categories within a subject that describe their properties and the relations between them. Essentially, ontology maps the terrain of a subject.
In science, it is essential to map the sexed attributes of an organism. Such should include all the significant waypoints that define an organism’s reproductive capacity. This is an entirely reasonable and scientific way of understanding human reproduction. In this way, “biological sex” does not exist in material reality in the precise way that cartography does not exist in material reality. Bodies and reproduction exist in material reality in the precise way that terrain and geological processes exist in material reality.
January 20, 2025, Trump signed Executive Order 14168, which asserts, “There are only two biological sexes,” defining “Female as the sex that produces the large reproductive cell (ovum), and male as the sex that produces the small reproductive cell (sperm).” Here, Trump has asserted that sperm production is what defines male. While this elementary school-level understanding of “biological sex” might make sense to the cult of MAGA, problematically, and as a matter of actual science, cis boys do not produce sperm for the first decade of their life.
To speak of “biological sex” in pop media is to speak of an imagined body binary wherein bodies come with only one of two discrete sets of sexed attributes. Trump communicates at a 3rd-grade level, and when he uses this term, he is appealing to elementary school ontology, and not anything as complex as the actual science of sex. When pop influencers take to (unintentionally ironic) “intellectual dark web” platforms to laugh at how dumb the left is because we don’t know that sex is only as complex as elementary school biology portrays, they’re demonstrating the Dunning-Kruger effect.
People who do not or cannot grasp that the world around them and their thoughts about the world around them are not the same things are people who, I would argue, fall into one of three categories: 1.) those who are not very bright; 2.) those who are profoundly confused about reality; or, 3.) those who are firmly on the road to sociopathy.
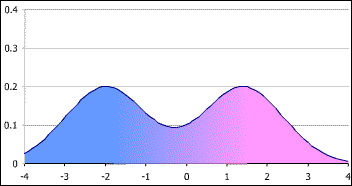
In very, very broad strokes there are around 50(ish) body attributes that present in material reality as a bimodal distribution spectrum. These spectrums include:
- Genetic Attributes:
- Sexed genes patterns: presence/absence/function of SOX9, DAX1, WNT4, FOXL2, SRY, SF1/NR5A1, WT1, DAX1/NR0B1, RSPO1, WNT4, DMRT1, GATA4, AMH/MIS, AMHR2, FGF9, DHH, CYP11A1, CYP17A1, HSD17B3, STAR, LHCGR, FSHR, AR, ESR1/ESR2, PGR, CYP19A1, HSD17B1, SRD5A2, LHβ/FSHβ, GNRH1, KISS1/KISS1R, HOXA10, HOXA11, HOXA13, LHX9, EMX2, PBX1, the Müllerian-inhibiting genes, FGF8, FGF10, BMP4, BMP7, SHH, ESR1, ESR2, CYP19A1, GNRH1, KISS1, TAC3, TACR3, IGF1, and NR3C1 genes.
- Gonadal Attributes:
- Ovarian tissue patterns: follicles, ova, etc.
- Testicular tissue patterns: seminiferous tubules, Leydig and Sertoli cells, etc.
- Ovotestes patterns: mixed gonadal tissue in some intersex variations.
- Hormonal Attributes:
- Androgen level patterns: testosterone, dihydrotestosterone, etc.
- Estrogen level patterns: estradiol, estrone, etc.
- Progesterone level patterns.
- Anti-Müllerian hormone level patterns.
- Gonadotropins (LH, FSH) level patterns.
- External Genitalia:
- Phallus: penis or clitoris (size, morphology).
- Labia majora/scrotum.
- Labia minora.
- Urethral position (perineal → penile).
- Vaginal opening.
- Post-Puberty Secondary Sex Characteristics:
- Breast development: glandular tissue.
- Facial hair/body hair distribution.
- Voice pitch via laryngeal growth.
- Wider/narrower pelvic morphology.
- Fat distribution: hips vs. abdomen.
- Muscle mass/strength distribution.
- Adams apple prominence.
- Skin texture and oiliness.
- Internal Reproductive Anatomy:
- Müllerian derivatives: uterus, fallopian tubes, cervix, upper vagina.
- Wolffian derivatives: epididymis, vas deferens, seminal vesicles.
- Glands: prostate-specific antigen secreting gland, bulbourethral/bartholin gland.
- Developmental/Physiological Attributes
- Menstrual cycling/ovulation.
- Spermatogenesis.
- Capacity for gestation.
- Ejaculatory function.
- fecundity markers.
- Biological Brain and Behavioral Correlates:
- Sex hormone receptor distribution in brain regions (hypothalamus, amygdala, etc.).
- Prenatal androgen exposure markers like digit ratio 2D:4D.
- Statistical sex differences in hypothalamic nuclei size.
Taken together, these attributes present in each human body as a bimodal distribution. It is simply scientifically false to assert that “sex” presents as a strict body binary.
And yes, I know the above list left out chromosomes. Because there are just XX and XY, right? In material reality, there’s a lot more variation than just that:
| Chromosome | Key Genes | Function in Sex Development | Variation | Phenotypic Effects |
|---|---|---|---|---|
| Y (Yp11.3, Yq11) | SRY, AZF (DAZ, RBMY, USP9Y) | SRY: Testis determination; AZF: spermatogenesis | 45,X/46,XY mosaic, Y microdeletions, Swyer syndrome | Ambiguous genitalia, gonadal dysgenesis, male infertility |
| X (Xp, Xq) | AR, DAX1 (NR0B1), FOXL2, SOX3, AMELX | AR: Androgen signaling; DAX1: Ovarian development; FOXL2: Ovarian maintenance | Turner (45,X), Triple X (47,XXX), Klinefelter (47,XXY), Fragile X, Ring X | Hypogonadism, ovarian failure, gonadal dysgenesis, infertility |
| 1p36 | NR0B2, LHX9 | Gonad formation | 1p36 deletion syndrome | Ambiguous genitalia |
| 2p23, 2q31 | LHCGR, HOXD cluster | LH receptor; external genitalia formation | LHCGR variations | Pseudo-hermaphroditism, Leydig cell hypoplasia |
| 3q23 | FOXL2 | Ovarian granulosa cell function | BPES syndrome (Blepharophimosis, ovarian failure) | Premature ovarian failure, sex reversal |
| 6q25 | ESR1 | Estrogen receptor α | ESR1 variations | Affects puberty, breast development |
| 7q11.23 | ELN, LIMK1 (Williams region) | Developmental/behavioral | Williams duplication / deletion | Variable effects, some genital anomalies |
| 7p15.3 | HOXA cluster (HOXA10, HOXA11, HOXA13) | Patterning of uterus, cervix, external genitalia | HOXA13 variations (Hand–Foot–Genital syndrome) | Urogenital tract malformations |
| 9q33.3 | DMRT1 | Testis development, Sertoli cell fate | 9p deletion (sex reversal) | XY sex reversal, gonadal dysgenesis |
| 10q26 | FGFR2 | Gonadal ridge, external genitalia | FGFR2 variations | Disorders of sexual development (rare) |
| 11p13, 11p15 | WT1, IGF2 | Gonadal ridge, kidney development | Denys-Drash, Frasier syndrome | Gonadal dysgenesis, nephropathy |
| 11q22-23 | ARSD | Steroid metabolism | Rare variants | Affects androgen metabolism |
| 11q13 | CCND1, NR5A1 (SF1) | Steroidogenic factor, adrenal/gonadal development | SF1 variations | 46,XY DSD, adrenal failure |
| 12p13.31 | HOXC cluster | Genital patterning | HOXC variations | External genital variations |
| 15q11-q13 | UBE3A, SNRPN | Genomic imprinting, hypothalamus | Prader–Willi, Angelman | Hypogonadism, delayed puberty |
| 17q12 | HNF1B | Urogenital development | 17q12 deletion syndrome | Renal/urogenital variations |
| 17q21.1 | SOX9 | Sertoli cell differentiation, testis cord | SOX9 duplication/deletion | Campomelic dysplasia, XY sex reversal |
| 17q24.2 | SRD5A2 | Conversion of testosterone → DHT | 5α-reductase deficiency | Undervirilized variations external genitalia |
| 19p13.2 | AMH | Regression of Müllerian ducts | AMH variation | Persistent Müllerian duct syndrome |
| 19p13.3 | AMHR2 | AMH receptor | AMHR2 variation | Persistent Müllerian duct syndrome |
| 19q13.33 | CYP11A1 | Initiates steroidogenesis | CYP11A1 variation | Congenital adrenal insufficiency |
| 19q13.32 | CYP19A1 (Aromatase) | Conversion of androgens → estrogens | Aromatase deficiency/excess | Virilization, ambiguous genitalia |
| 19q13.12 | HSD17B3 | Testosterone synthesis | HSD17B3 deficiency | Under/un-masculinization, XY DSD |
| 20q13.2 | BMP2, BMP7 | External genitalia morphogenesis | BMP variations | Hypospadias, ambiguous genitalia |
| 22q11.2 | TBX1 | Reproductive tract patterning | DiGeorge syndrome | Cryptorchidism, urogenital anomalies |
An elementary school education often simplifies the science of sex down to the notion that the presence or absence of a Y chromosome in a human body will determine sex because only men have bodies with Y chromosomes. In material reality, that’s not factual. Almost all female bodies on the planet have Y chromosomes because most were born with them. Applying an elementary school understanding of sex, this means that most women on the planet are men because only men have Y chromosomes.
The perfectly natural condition is called microchimerism. It means that the bodies of women mostly have XX chromosomes, but fewer XY chromosomes, and the bodies of men mostly have XY chromosomes, with fewer XX chromosomes. Most people alive today have this condition. If you are a cis man, you probably have a body with some XX chromosomes; that doesn’t make you a woman. If you are a cis woman, you probably have a body with some XY chromosomes; that doesn’t make you a man.
A popular claim for those whose sex education never made it out of elementary school will be that somehow “every cell” in your body is sexed and that scientists can determine the sex of a trans or intersex person from their bones. For instance, this right-wing hate group makes this very claim to justify their anti-trans and anti-intersex bias:
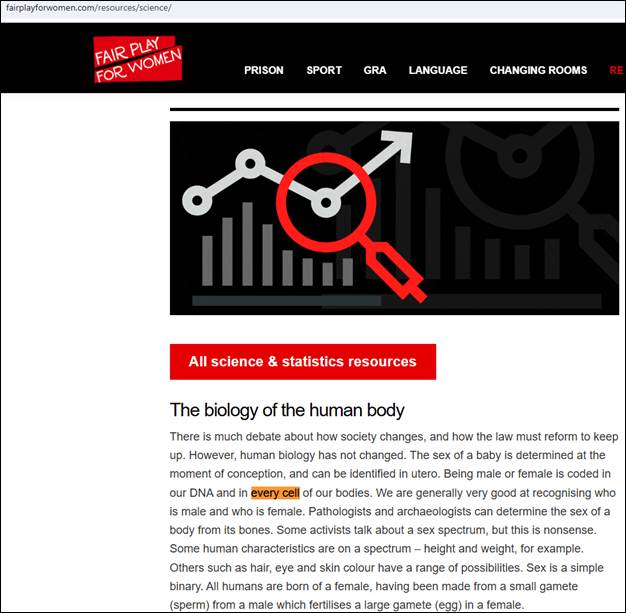
Here, the hate group is appealing to the chromosomal DNA in cells. Human erythrocyte, thrombocyte, keratinocyte, lens fiber cells, and, unintentionally ironic to the hate group’s point, spermatozoa (which have no cellular chromosomal DNA) are not, by their own definition of sex, sexed cells. None of these human cells contain their own chromosomal DNA.
This says nothing about the scientific fact that medical transition changes DNA form, which is to say, one’s genotype form. Let me repeat that: it is a scientific fact that medical transition changes DNA form and DNA expression. Trans and intersex people who undertake medical transition do not have the stereotypical DNA of the sex assigned to them at birth. DNA methylation is the chemical addition of methyl groups to DNA, serving as an epigenetic “switch” that regulates gene expression, and it is this sexed attribute that is transitioned during medical transition. Specifically, there are at least 5,657 differentially methylated CpG sites (DMPs) that become female-typical for trans women during medical transition. A CpG site is a place in DNA where a cytosine (C) nucleotide is directly followed by a guanine (G) nucleotide. DNA is comprised of adenine (A), guanine (G), cytosine (C), or thymine (T) nucleotides.

In other words, when people with an elementary school sex education confidently assert that transition doesn’t “change your DNA,” they’re demonstrating their scientific ignorance. As an aside, I wonder what a Venn diagram might look like for the people who claim that transition doesn’t change one’s DNA and those who were very concerned that their COVID-19 vaccination would change their human DNA into non-human DNA.
Regarding bones, even for the cisgender population, sex determination is not accurate. For example, for cisgender men, there’s about a 30% chance that their long bones will not be sexed as male. The notion that transgender and intersex medical transition has no impact on bone development is not consistent with reality. Particularly, for trans youth who undergo puberty blockers followed by medical transition, their bones tend to resemble those of their affirmed gender. Specifically, van der Loos et al., 2021 found that trans adolescents treated with puberty blockers followed by gender-affirming HRT (GAHT) had bone density and geometry consistent with their affirmed gender; Schagen et al., 2020 and Klink et al., 2015 found that GAHT patterns the bones of trans kids to that of their cis peers of the same affirmed gender.
Lastly, I feel it important to state that there is an implicit genocidal aspect to pop culture’s “biological sex” dialectic. To assert that trans and intersex people are not biological in the way that cisgender people are can lead to some very dark places wherein we as a society begin to regard trans and intersex people as something synthetic or inhuman. Sex is genotype and phenotype; the transitioned phenotype and genotype of trans and intersex people are as biological as any cis person’s genotype and phenotype. The transitioned sexed attributes of a trans or intersex person’s body are biological. It doesn’t matter if a child went through a medical puberty or a natal puberty; their resulting sexed attributes are just as biological as any cis person’s sexed attributes are. If you find yourself questioning whether trans and intersex people are just as biological as cis people, you need to do some serious self-reflection because you’re on the road to functional sociopathy.
In sum: Bodies and reproduction exist in material reality; your thoughts about that reality do not.
- Elementary school biology is not science.
- Pop culture biology is not science.
- Thoughts about bodies and reproduction are not the same thing as bodies and reproduction.
- Thoughts about a body’s sexed attributes are gender.
- All people, including trans and intersex people, are biological.